
Since de novo lipogenesis (the conversion of carbs to fat) is quantitatively unimportant in humans, how does the body get rid of the excess calories from carbs?
Let’s take a closer look at carbohydrate disposal. After glycogen depletion, glycogen stores takes up 4 days to saturate (1). Extreme carbohydrate manipulations have shown rapidly auto-regulatory adjustments in carbohydrate oxidation rates (2,3) over short periods of time, and the effect persists after normalization of the diet in response to the perturbed glycogen stores (2).
Let’s take a closer look at carbohydrate disposal. After glycogen depletion, glycogen stores takes up 4 days to saturate (1). Extreme carbohydrate manipulations have shown rapidly auto-regulatory adjustments in carbohydrate oxidation rates (2,3) over short periods of time, and the effect persists after normalization of the diet in response to the perturbed glycogen stores (2).
By the end of the second day of overfeeding, glycogen stores had increased by 500g. At this point carbohydrate oxidation and storage became insufficient to dispose of all of the ingested carbohydrate, and some had to be converted to fat, ie, de novo lipogenesis (150g lipid/d using 475 CHO/d). This is perhaps the biggest increase in DNL ever recorded, possibly due the extreme nature of the diet with only 3% of fat and with 86% of carbohydrate. It is known that DNL increases a bit more with a diet below 10% of fats coupled with a high carb intake.
After 4 d of overfeeding glycogen stores became saturated at ~770g (occurred on day 5 for one subject). When the glycogen stores are saturated, massive intakes of carbohydrate are disposed of by high carbohydrate-oxidation rates (1).
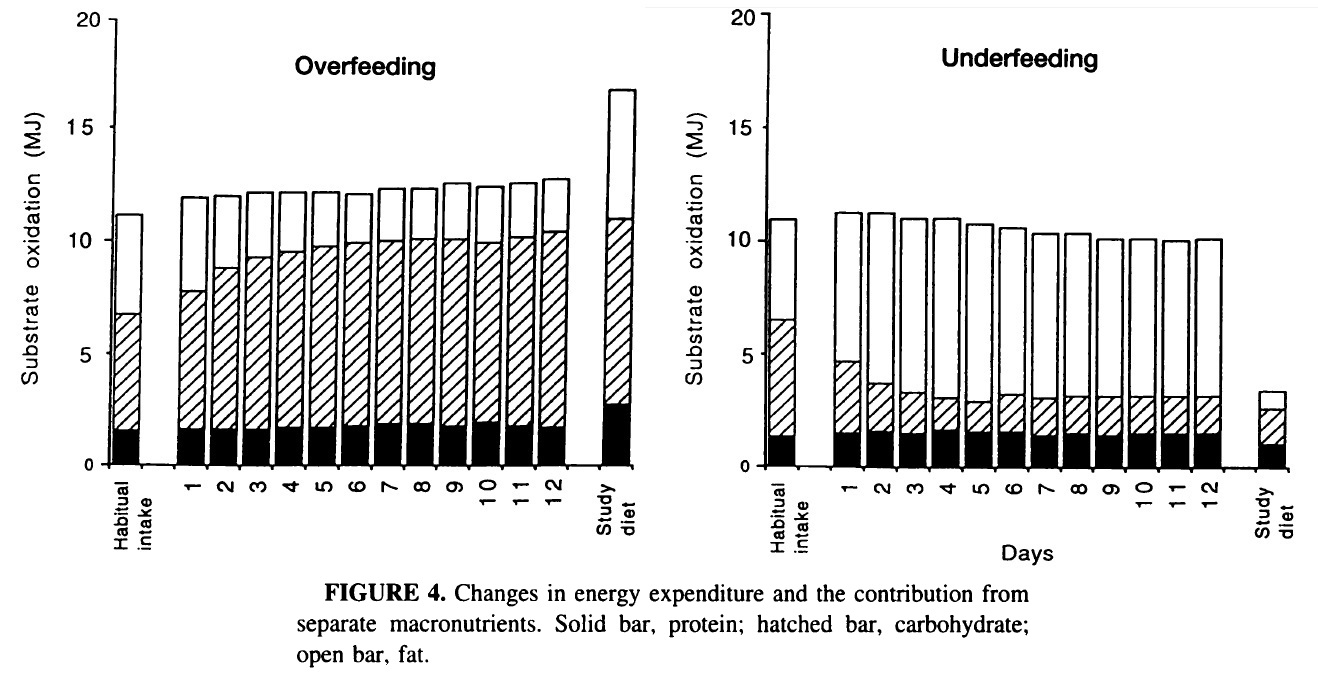
In other metabolic ward study (12 days),
carbohydrate intake of 540g and 83 g/d for overfeeding and underfeeding,
respectively, exerted direct auto-regulatory feedback on carbohydrate oxidation
(551 and 106 g/d at day 12 for overfeeding and underfeeding, respectively) (4).
With carbohydrate overfeeding there was a large
increase in carbohydrate oxidation but also in glycogen storage (339 ± 82
g/d). Carbohydrate balance was achieved after the first few days and by day 12
carbohydrate oxidation was 551 g/d compared with an intake of 539 g/d.
Carbohydrate oxidation was providing ~8.68 MJ/d, thus fat oxidation was
suppressed. During overfeeding, BMR increased by 0.42 MJ (5.7%) and TEE
increased by 0.75 MJ (6.2%).
This study also performed underfeeding.
As expected over the first few days of underfeeding there was a sharp decrease
in carbohydrate oxidation, reflecting a gradual but progressive decrease in
muscle glycogen. After day 4 carbohydrate intake and oxidation were closely
matched, with a small, persistent daily negative carbohydrate balance with
intakes of 83 g/d compared with oxidation of 106 g/d (1.67 MJ/d) on day 12. In
this case, to meet the body’s energy requirement endogenous fat oxidation
increased. BMR decreased by 0.82 MJ (8.3%) and TEE decreased by 1.20
MJ (10.5%).
The contribution of protein to the fuel mixture
during both interventions remained remarkably constant.
 (4)
(4)
The increased energy expenditure was approximately double that which could be explained by the combination of increased TEF and increased body mass, meaning that more of the excess energy was oxidized and less stored in the body than was seen during fat overfeeding (5).
Even on day 14 total energy expenditure was higher with carbohydrate overfeeding so that the total stored energy was less than with fat overfeeding.

This study also compared lean vs. obese subjects, and
it was noted that obese
subjects oxidized proportionally more carbohydrate and less fat than did lean
subjects (5). The greatest reliance on carbohydrate oxidation during energy
balance perturbations may be a risk factor for obesity (6,7,8). Subjects with the
highest oxidative capacity of skeletal muscle have the lowest ratio of fat to
lean mass in weight gain (9).
Another study showed a graded dose response in
carbohydrate oxidation (10). Researchers used stable isotope-mass spectrometric
methods with indirect calorimetry in normal subjects to quantify the metabolic
response to six dietary phases (5 d each), ranging from 50% surplus CHO (+50%
CHO) to 50% deficient CHO (-50% CHO), and 50% surplus fat (+50% fat).
A dose response was observed in glucose production with increasing carbohydrate intake, which stimulated moderate hyperinsulinemia and decreased lipolysis and fatty acid availability. The net effect was to increase glycogen stores and deliver extracellular glucose, thus favoring increased carbohydrate oxidation and a reciprocal decrease in fat oxidation (10).
A dose response was observed in glucose production with increasing carbohydrate intake, which stimulated moderate hyperinsulinemia and decreased lipolysis and fatty acid availability. The net effect was to increase glycogen stores and deliver extracellular glucose, thus favoring increased carbohydrate oxidation and a reciprocal decrease in fat oxidation (10).
Carbohydrate disposal is not different between
glucose, fructose, and sucrose, in lean and obese women (11). Different
carbohydrates behave in an essentially identical manner. A 50% overfeeding
with either glucose,
fructose and sucrose resulted in no significant difference in fat balance,
and there were no significant differences between lean and obese women in
macronutrient oxidation or balances. As expected, carbohydrate oxidation
increased greatly in response to carbohydrate overfeeding (from 15.61 to 21.94,
21.64, and 21.97 MJ for fructose, glucose, and sucrose, respectively).
Of the excess carbohydrate, +74 % was oxidized (compared to only 18% of the excess fat intake) and on average 12%
of the excess energy was stored as glycogen and 88% as fat; there was no
significant difference between overfeeding treatments.
As seen in other studies (4,12), almost all of the
glycogen storage occurred on day 1, with minimal imbalance on subsequent
days, which may suggest the need for glycogen stores to first be perturbed to
generate feedback control.
The daily carbohydrate imbalance with sucrose overfeeding
asymptotically approached zero as carbohydrate oxidation gradually increased
until it exactly matched intake. This caused glycogen storage to plateau at a
new level ~110 g above the initial value.
Glycogen must be regulated within a relatively narrow
window, and adipose tissue has evolved as the main energy storage compartment.
Once any short-term changes in glycogen have resolved, additional energy excess
or positive imbalances are buffered by fat stores.
Carbohydrate vs. Protein
It was once thought that the auto-regulatory control
of protein oxidation was as efficient as that of carbohydrate (13). However, a
160% increase in protein intake (from 47 g/d during underfeeding to 122 g/d
during overfeeding) only caused a 12% increase in protein oxidation (from 83
g/d during underfeeding to 93 g/d during overfeeding) (4).
In contrast, a 550% increase in carbohydrate intake (from 83 g/d during underfeeding to 539 g/d during overfeeding) was almost matched by a 420% increase in oxidation (from 106 g/d during underfeeding to 551 g/d during overfeeding). Carbohydrate was much more responsive, and it certainly exerted a much greater influence on the reciprocal changes in fat utilization than did protein (4). Protein takes a subordinate position to carbohydrate in terms of oxidation.
In contrast, a 550% increase in carbohydrate intake (from 83 g/d during underfeeding to 539 g/d during overfeeding) was almost matched by a 420% increase in oxidation (from 106 g/d during underfeeding to 551 g/d during overfeeding). Carbohydrate was much more responsive, and it certainly exerted a much greater influence on the reciprocal changes in fat utilization than did protein (4). Protein takes a subordinate position to carbohydrate in terms of oxidation.
Would you like to know more? Subscribe for daily updates!
References:
1. Acheson KJ,
Schutz Y, Bessard T, Anantharaman K, Flatt JP, Jéquier E. Glycogen storage capacity and de novo
lipogenesis during massive carbohydrate overfeeding in man. Am J Clin Nutr 1988;48:240-7.
2. Shetty PS,
Prentice AM, Goldberg GR, et al. Alterations in fuel selection and voluntary
food intake in response to isoenergetic manipulation of glycogen stores in
humans. Am i Clin Nutr l994;60:534-43.
3. Schutz Y, Tremblay A, Weinsier RL, Nelson KM. Role
of fat oxidation in the long-term stabilization of body weight in obese women.
Am J Clin Nutr 1992;55:670-4
4. Jebb SA, Prentice AM, Goldberg GR, Murgatroyd PR,
Black AE, Coward WA. Changes in macronutrient balance during over- and
underfeeding assessed by 12-d continuous whole-body calorimetry. Am J Clin Nutr
1996;64:259–66
5. Tracy J Horton, Holly Drougas, Amy Brachey,
George W Reed, John C Peters, and James
Hill .Fat and
carbohydrate overfeeding in humans: different effects on energy storage. Am J Clin Nutr 1995;62:19-29.
6. Hill JO, Pagliassotti
MJ, Peters JC. Nongenetic determinants of obesity and fat topography. In:
Bouchard C, ed. Genetic determinants of obesity. Boca Raton, FL: CRC Press, Inc
1994:35-48.
7. Thomas CD,
Peters JC, Reed OW, Abumrad NN, Sun M, Hill JO. Nutrient balance and energy
expenditure during ad libitum feeding of high-fat and high-carbohydrate diets
in humans. Am J Clin Nutr 1992;55:934-42.
8. Zurlo F, Lillioja 5, Esposito-Del
Puente A, et al. Low ratio of fat to carbohydrate oxidation as
predictor of weight gain: study of 24-h RQ. Am i Physiol 1990;259:E650-7.
9. Dériaz O,
Fournier G, Tremblay A, Després JP, Bouchard C. Lean body-mass composition and
resting energy expenditure before and after long-term overfeeding. Am I Clin
Nutr 1992;56:840-7.
10. Schwarz JM,
Neese RA, Turner 5, Dare D, Hellerstein MK. Short term alterations in
carbohydrate energy intake in humans. I Clin Invest 1995;96:2735-43.
11. Regina M McDevitt, Sally D
Poppitt, Peter R Murgatroyd, and Andrew M Prentice. Macronutrient disposal
during controlled overfeeding with glucose, fructose, sucrose, or fat in lean
and obese women. Am J Clin Nutr 2000;72:369–77.
12. Schutz Y,
Acheson KJ, Jequier E. Twenty-four hour energy expenditure and thermogenesis:
response to progressive carbohydrate overfeeding in man. Int J Obes
1984;9:111–4.
13. Krebs HA.
Some aspects of the regulation of fuel supply in omnivorous animals. Adv Enzyme
Regul 1972;l0:397-420.